
Le phénotype blanc sourd a été rapporté chez de multiples mammifères, dont la souris, le chien, le vison, le cheval, le rat, le hamster doré, l'alpaga et l'homme. Chez le chat blanc sourd, la surdité se produit naturellement. Elle une base génétique et présente une expression variable. Chez l''humain, le syndrome de Waardenburg de type 2 représente le plus précisément le phénotype, avec une hypopigmentation caractéristique de la peau et des cheveux et une dysplasie cochléo-sacculaire qui ressemble à la malformation de Scheibe.
La surdité est à pénétrance incomplète, ce qui a empêché les précédentes tentatives de cartographie génétique. En outre, de multiples gènes situés sur de multiples loci interagissent sur l'audition : un gène unique est rarement responsable du fonctionnement d'un système.

Une étude publiée en 2014, portant sur 270 chats de 30 races différentes, a montré que les chats à panachure blanche portent une insertion d'un rétrovirus endogène dans une zone précise de KIT. Les chats entièrement blancs portent une version très réduite du même rétrovirus, qui est clairement associé avec la surdité. Les rétrovirus endogènes sont des copies de rétrovirus exogènes insérés dans le génome d'un hôte via des infections anciennes. En se fondant sur le génotype des chats étudiés, les chercheurs ont montré qu'il existe une corrélation entre W et la probabilité qu'un individu soit sourd et/ou aux yeux bleus et il y a une corrélation indéniable entre l'homozygotie pour W et la surdité des chats blancs. Cette corrélation ne signifie cependant pas que la relation est causale.
Une ou 2 copies de l'insertion partielle du rétrovirus causent le phénotype blanc dominant (DW), donc rendent le chat blanc quel que soit l'autre allèle, mais on observe des degrés variables de déficience auditive. Un chat dont les 2 yeux sont bleus a 3 à 5 fois plus de risque d'être sourd qu'un chat aux yeux non bleus. Un chat avec un œil bleu a environ 2 fois plus de risque d'être sourd qu'un chat aux 2 yeux non bleus. De plus, les chats blancs à poil long ont 3 fois plus de risque de surdité bilatérale. A l'état sauvage, les chats blancs sourds subissent une forte pression négative de sélection naturelle car :

En étudiant un pedigree étendu, des chercheurs ont constaté que presque 75% des chats avec 2 copies de DW étaient sourds et que les non sourds avaient au moins une déficience auditive partielle. Chez les chats avec une seule copie de DW et une copie normale pour l'autre allèle, près de 60% des chats avaient une audition normale, 21% étaient sourds et 17% avaient une déficience auditive. Chez les chats avec une copie de DW et une copie de Ws, 1/3 avaient une audition normale, 1/3 avaient une déficience auditive et 1/3 étaient sourds.
Une autre étude sur la prévalence de la surdité sur la base du test de BAER a été effectuée chez 84 chats blancs de 10 races pures. Les chercheurs ont trouvé des taux de prévalence similaires : 9,5% de chats sourds unilatéralement et 10,7% bilatéralement, soit 20,2% des chats touchés par une surdité. On a observé des chats sourds chez 6 des 10 races : Angora Turc, British Shorthair, Maine Coon, Norvégien, Persan et Foreign White. Aucune différence n'a été constatée entre les sexes mais les chats aux yeux bleus étaient plus à risque pour la surdité que ceux aux yeux non bleus.
Au début de l'embryogénèse, les mélanoblastes migrent de la crête neurale vers la peau, les régions de l'œil et celles de l'oreille interne. Les mutations qui affectent toutes les étapes de ce parcours, que ce soit la prolifération, la survie, la migration ou la distribution des mélanoblastes se manifestent souvent par une variation de la couleur de la fourrure. On a décrit chez des mutants de souris à panachure de nombreux gènes, dont KIT, qui ont été identifiés dans ces éléments précoces de pigmentation.
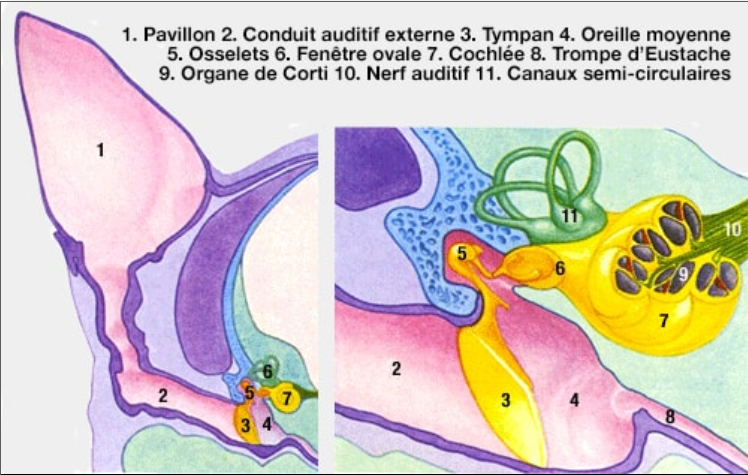
On peut subdiviser les mélanocytes en lignées cutanées et non cutanées, qui répondent différemment aux signaux de KIT pendant le développement. Les mélanocytes qui colorent la peau et les poils sont très sensibles aux signaux de KIT, alors que les mélanocytes de l'oreille interne et des régions de l'œil, de l'iris et de la choroïde, sont stimulés plus efficacement par d'autres facteurs. Dans l'oreille interne, on observe de nombreux mélanocytes dans la strie vasculaire, l'épithélium vascularisé étant responsable de la sécrétion de niveaux élevés de potassium vers l'endolymphe, qui établit le potentiel endocochléaire (PE). Le potentiel endocochléaire est indispensable au fonctionnement normal des cellules des récepteurs auditifs. Les mélanocytes sont nécessaires au fonctionnement du PE, donc leur absence cause une surdité.
On pense que la pénétrance incomplète de la couleur de l'iris et de la surdité résulte d'une combinaison de facteurs génétiques, environnementaux et de mode de vie, dont beaucoup sont inconnus. Le manque de pénétrance apparente de la surdité sur W peut être expliqué si KIT est identifié comme le locus félin du blanc. Chez les individus hétérozygotes pour la mutation, certains mélanocytes pourraient survivre à la migration vers l'oreille interne et l'iris car ils sont moins sensibles à une diminution du signal de KIT, contrairement aux mélanocytes destinés à pigmenter les poils, qui sont extrêmement sensibles à l'expression de KIT. L'achèvement de l'analyse de liaison pourrait fournir la réponse à cette question de pénétrance.
Surdité et pigment
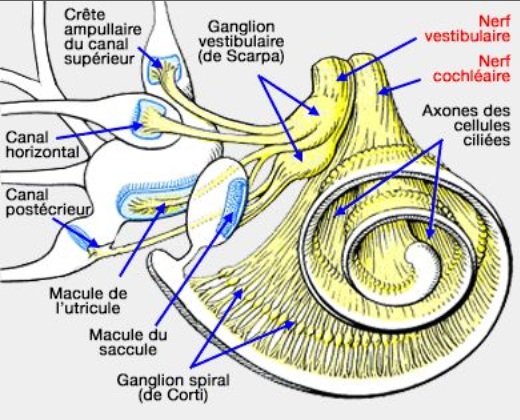
La plupart des surdités héréditaires chez le chat relèvent de la pathologie cochléo-sacculaire, aussi nommée pathologie de type Scheibe, qui est connue chez le chat blanc. Elle résulte d'une dégénérescence primaire de la strie vasculaire sur la paroi externe du canal cochléaire. L'expression est le plus souvent unilatérale mais peut toucher les 2 oreilles. Aucun potentiel endocochléaire ne subsiste. La dégénérescence striale initiale est généralement liée à l'absence de mélanocytes fonctionnels. On peut donc dire qu'elle est associée au pigment. Cette surdité reproduit la dysplasie de Scheibe chez l'humain, une manifestation post-natale précoce d'un déficit auditif neurosensoriel transmis via un modèle autosomique dominant à pénétrance incomplète.
Blanc ou albinos ?
Les chats blancs ne sont pas nécessairement albinos.
Anomalies anatomiques liées à la surdité
L'apparence de l'oreille interne des chats sourds est nettement différente de celle des chats entendants. La surdité est révélée par une incapacité à provoquer une réponse du cerveau à un son et est couplée à une pathologie de la cochlée.
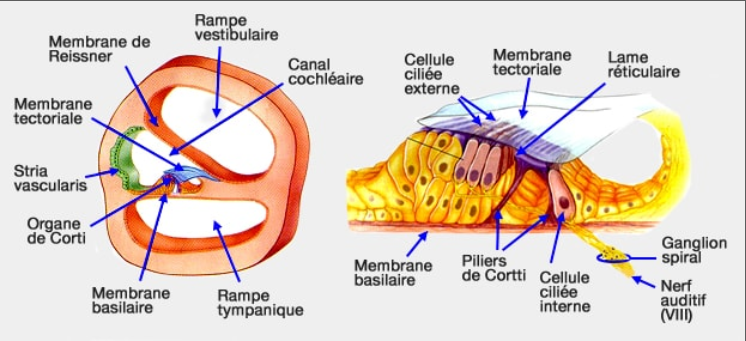
La surdité chez les chats blancs est corrélée à 2 types d'anomalies structurales :
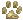 Dans la forme la plus fréquente, la membrane de
Reissner est descendue sur
l'organe de
Corti et le canal cochléaire est obturé. L'obturation se produit pendant les
10 jours qui suivent la naissance. La strie vasculaire est présente, mais nettement plus
mince
que la normale. Après la 3e semaine après la naissance, le canal de
l'oreille externe s'ouvre, les chatons
qui
à
cet âge ne
répondent absolument pas à la stimulation acoustique n'ont pas de canal
cochléaire. Chez les chats
sourds plus âgés, l'organe de Corti est quasiment impossible à
reconnaître.
Dans la forme la plus fréquente, la membrane de
Reissner est descendue sur
l'organe de
Corti et le canal cochléaire est obturé. L'obturation se produit pendant les
10 jours qui suivent la naissance. La strie vasculaire est présente, mais nettement plus
mince
que la normale. Après la 3e semaine après la naissance, le canal de
l'oreille externe s'ouvre, les chatons
qui
à
cet âge ne
répondent absolument pas à la stimulation acoustique n'ont pas de canal
cochléaire. Chez les chats
sourds plus âgés, l'organe de Corti est quasiment impossible à
reconnaître.
Dans l'autre forme de pathologie cochléaire,
on constate une prolifération
de
cellules tout au long de la spirale cochléaire. La membrane de Reissner est
hypertrophiée, devient très irrégulière et pliée, et finit
par
remplir le canal cochléaire. Les cellules de soutènement
de l'organe de Corti
et les cellules épithéliales de la membrane basilaire sont hypertrophiées.
La
membrane basilaire est
déformée et le canal de Corti n'atteint pas sa forme triangulaire
caractéristique, les cellules
ciliées ne se sont pas différenciées et la raie vasculaire est
voilée.
Globalement, les tissus
présentent une apparence « spongiforme ». Le corps cellulaire des neurones du
ganglion spiral est
bien dans le système auditif périphérique mais ces neurones étendent
leurs terminaisons centrales
dans le système auditif central. La survie de ces neurones ganglionnaires dépend
de la
santé de
l'organe de Corti car ils subissent une dégénérescence associée
à
la perte de cellules ciliées et la
surdité neurosensorielle. Chez les chats atteints par une surdité
congénitale,
il y a une perte
graduelle de cellules du ganglion spiral avec l'âge. (les neurones
bipolaires
du
ganglion
spiral sont connectés aux cellules ciliées par une de leurs fibres, l'autre,
l'axone, forme le nerf cochléaire).
Il y a plus de 140 ans, Charles Darwin a remarqué qu'en comparaison avec leurs ancêtres sauvages les espèces domestiques sont plus dociles. Il a également noté qu'elles tendent aussi à présenter un ensemble d'autres caractères spécifiques, notamment des oreilles repliées, des faces plus juvéniles avec des mâchoires plus petites et... des taches de fourrure blanche.

Bien plus tard Dimitri Beljaev, un généticien soviétique, a émis l'hypothèse que le phénotype exprimé par les animaux domestiques ne résultait pas d'une sélection délibérée sur des caractères individuels, mais qu'il était la conséquence de la sélection en vue d'une caractéristique comportementale. En 1959, il a commencé à tester son hypothèse en élevant des renards argentés, des animaux jamais domestiqués auparavant. Son critère de sélection : l'agressivité la plus faible possible. Beljaev espérait qu'en sélectionnant un caractère comportemental spécifique, il influencerait le phénotype des générations suivantes en le rendant d'apparence plus domestique. L'expérience fut un succès : outre une disposition amicale envers les humains, les renards finirent par présenter des caractères qui distinguent en général les animaux domestiques de leur ancêtre sauvage, comme les oreilles pendantes, la queue recourbée, le museau plus court etc.
Outre démontrer la facilité d'apparition de caractères phénotypiques domestiques via une sélection pour un caractère comportemental, Beljaev a fourni un mécanisme qui explique comment la domestication précoce des animaux pourrait avoir commencé sans action humaine délibérée. Beljaev avait effectué sa sélection uniquement en vue de la docilité, et vu l'absence d'autres choix sélectifs au cours des premières étapes de son travail, il était concevable que les gènes contrôlant le comportement (tout au moins la docilité) étaient aussi impliqués dans l'apparition des fourrures tachetées de blanc. Les renards de Beljaev ont montré que la panachure pouvait apparaître après seulement 10 générations d'élevage. Cela a rendu plausible l'idée qu'une nouvelle variation peut se produire rapidement, et qu'aussi longtemps que les humains contrôleront la reproduction de leurs animaux domestiques, ils encourageront la multiplication des variantes de couleurs de fourrure. La fréquence des nouveaux phénotypes augmente jusqu'à ce que de nouvelles mutations se produisent, menant à une variation plus importante et une distanciation moléculaire plus importante par rapport à l'allèle sauvage.
Le fait que certains gènes sont responsables des phénotypes domestiques suggère aussi que la variation apparaît tôt, quel que soit le chemin que l'animal a suivi vers la domestication.
Et la crête neurale ?

Aujourd'hui, Adam Wilkins, Richard Wrangham et Tecumseh Fitch cherchent à expliquer pourquoi les animaux domestiques ont tous en commun un ensemble de caractéristiques incluant la docilité, des faces courtes, des oreilles pendantes et des taches de fourrure blanche. Leur hypothèse pose que les mammifères domestiqués peuvent présenter un déficit du développement ou de la migration des cellules de la crête neurale par rapport à leurs ancêtres sauvages. « Quand les humains ont élevé ces animaux sur le critère de la docilité, ils ont pu par hasard sélectionner les individus avec de légères déficiences de la crête neurale, qui se traduisent par des glandes surrénales plus petites ou à croissance plus lente. Donc ces animaux étaient moins craintifs » dit Wilkins. Wilkins et al. (2014) suggèrent qu'une forte sélection orientée vers la docilité pourrait causer des modifications de la manière dont les glandes surrénales et le système nerveux sympathique se développent à partir de la crête neurale et que les effets collatéraux de ces déficits pourraient inclure diverses modifications physiques.
Entre autres effets, un déficit de la crête neurale peut causer la dépigmentation de certaines zones de la peau (p. ex. des taches blanches), un cartilage d'oreilles mal formé, des dents plus petites, et des changements dans le développement de la mâchoire, tous ces effets étant inclus dans le syndrome de domestication. Les auteurs suggèrent aussi que la taille réduite du cerveau antérieur de la plupart des mammifères domestiques pourrait être un effet indirect des modifications de la crête neurale. En effet, un signal chimique envoyé par ces cellules est indispensable à un développement correct du cerveau.
L'un des nombreux changements observés chez les renards apprivoisés a été la taille et la fonction réduite des glandes surrénales, qui émettent des hormones de stress pendant la réponse « combat-fuite ». Cette fonction réduite des surrénales est peut-être au cœur des changements de comportement observés lors du syndrome de domestication. Wilkins et al. considèrent qu'une des causes de la réduction des glandes surrénales est un déficit léger de la crête neurale. Nous avons vu plus haut que les cellules de la crête neurale migrent vers de nombreuses parties du corps et forment les précurseurs de nombreux types de tissus, dont les cellules pigmentaires, des parties du crâne, du larynx, des oreilles, des dents, du système nerveux sympathique et, bien sûr, des parties des glandes surrénales. Donc, des changements subtils de la crête neurale (nombre de cellules, migration ou prolifération) pourraient causer des effets phénotypiques étendus.

Wilkins et al. considèrent que leurs idées sont corroborées par certains effets des désordres de la crête neurale chez l'humain, comme des taches dépigmentées sur la peau et les cheveux dans le syndrome de Waardenburg ou les phénotypes de la mâchoire, de l'oreille et des dents dans le syndrome de Treacher Collins. Et, bien que les cellules de la crête neurale ne se transforment pas directement en système nerveux central, elles pourraient expliquer en partie pourquoi de nombreux animaux domestiques ont un cerveau antérieur plus petit que leurs ancêtres sauvages (des expériences suggèrent notamment que les signaux de la crête neurale jouent un rôle crucial dans le développement du cerveau antérieur). Les auteurs suggèrent aussi que certains loci associés à la docilité, comme ceux découverts chez les rats et les renards domestiques, sont impliqués dans la biologie de la crête neurale.
En identifiant les traces de la domestication grâce à la découverte de zones génétiques qui expriment les signatures de la sélection chez le chat domestique mais non chez ses parents sauvages, les auteurs de l'étude sur le génome du chat ont aussi apporté un soutien à l'hypothèse du syndrome de domestication lié à la crête neurale. En effet, ils suggèrent que le lien sous-jacent entre ces caractères pourrait être la crête neurale. Cette proposition n'a pas encore été vérifiée mais c'est la 1e hypothèse qui connecte plusieurs composantes du syndrome de domestication. A ce stade, on ne peut pas relier solidement toutes les composantes du syndrome de domestication à l'hypothèse : par exemple, la queue enroulée des chiens, des porcs et des renards domestiques n'a pas de lien évident avec les déficits de la crête neurale. Cependant, les auteurs croient qu'il existe assez de liens pour valider leur hypothèse.
Une possibilité est que de telles différences sont dues à de nombreux allèles aux effets relativement faibles sur le développement de la crête neurale plutôt qu'à quelques variations aux effets importants.
Cette idée fondée sur la biologie du développement fournit une hypothèse unificatrice et apporte un aperçu précieux de la biologie de la domestication. La vérification de l'hypothèse de la crête neurale est peut-être proche, car des scientifiques cartographient rapidement les gènes qui ont été altérés par la domestication chez le rat, le renard et le chien, et certains de ces gènes influeraient sur la biologie de la crête neurale. Si c'est le cas, nous comprendrons bien plus en profondeur la biologie sous-jacente à un événement significatif de l'évolution, dit Wilkins. « La domestication animale a été une étape cruciale du développement des civilisations humaines. (...) Sans les animaux, il est difficile dimaginer que les sociétés humaines auraient prospéré comme elles l'ont fait. »
L'hypothèse de la crête neurale pourrait-elle aussi expliquer pourquoi les allèles de panachure de KIT sont si fréquents chez les chats ? Les mutations de KIT affectent la migration d'une population issue de la crête neurale, mais ces mutations ne sont pas connues pour affecter le comportement. A moins qu'il n'ait un effet pléiotropique à la fois sur la pigmentation et sur la docilité, ce gène ne peut pas être un contributeur majeur au syndrome de domestication.
Presque 100 ans après que le Dr Whiting ait déduit l'existence des allèles contrôlant le blanc à partir de croisements tests, nous connaissons leur identité et leur séquence et pouvons commencer à déterminer leurs conséquences fonctionnelles et évolutives.

Illustrations :
Sources :
{kind=link}