Préalable
Nous avons choisi de vous offrir la traduction aussi exacte que possible de cette étude. Le texte fait référence aux méthodes qui ont été utilisées afin de réaliser cette étude basée sur l'analyse d'une partie du génome de nombreux chats. Il n'est pas dans notre idée (ni de notre niveau) de vous détailler ces techniques : Les conclusions de cette étude sont intéressantes en elles-mêmes. Nous avons cependant conservé, sous forme adaptée et simplifiée, quelques annexes auxquelles vous pouvez accéder via les ancres 1 à 7.
L’expansion des races de chats : évaluation génétique de races et de populations spontanées du monde entier
Résumé
La diaspora du chat moderne a été pistée avec des marqueurs microsatellites [1.] depuis le site de domestication présumé jusqu’à des régions éloignées du monde. Les données génétiques ont été obtenues à partir de plus de 1100 individus, représentant 17 populations spontanées de cinq continents et 22 races [2.]. La région méditerranéenne a été reconfirmée comme site de domestication probable. La diversité génétique est restée importante dans le monde entier, avec des groupes génétiques distincts dans le bassin méditerranéen, en Europe/Amérique, Asie et Afrique. Cependant, les chats asiatiques semblent s’être séparés tôt des autres et s’être étendus dans un relatif isolement. La plupart des races sont dérivées des chats indigènes de leur région d’origine supposée. Cependant, le Persan et le Bobtail japonais se sont avérés plus proches des groupes Européens/Américains qu’avec ceux du bassin méditerranéen ou d’Asie. Trois races récemment dérivées (crées) ne sont pas distinctes de leurs races parentes d’origine. La race pure a été associée à une perte de diversité génétique, cependant, cette perte n’est pas corrélée avec la popularité ou l’âge de la race.
Le chat domestique (Felis Catus, Linné 1758) est prolifique et cosmopolite ; il occupe la plupart des parties habitables du monde. Les données archéologiques suggèrent que la domestication du chat s’est produite au proche orient, il y a environ 9 à 10 000 ans. Cependant, le début de la domestication a probablement commencé des milliers d’années plus tôt, quand les humains et les chats ancestraux sont progressivement devenus plus interdépendants. Le processus de domestication a vraisemblablement commencé au cours de la période pendant laquelle les humains ont cessé de suivre les troupeaux d’animaux sauvages pour adopter des modes de vie plus agricoles, en particulier dans le Croissant Fertile.
Le Croissant fertile
C'est une région du Moyen-Orient comprenant les territoires des États actuels d' Israël, du Liban, de Chypre, du Koweït, de Palestine (Cisjordanie et Gaza) ainsi que des parties de la Jordanie, de la Syrie, de l'Irak, de l'Iran, de l'Égypte (sur ce point, il ne semble pas y avoir consensus) et le sud-est de la Turquie. Le terme « Croissant fertile » fut donné par l'archéologue James Henry Breasted de l'Université de Chicago car l'arc formé par les différentes zones ressemble à un croissant.Ce changement s’est produit il y a 10-11 000 ans, et a été rendu possible par la domestication de certaines herbes et graminées sauvages. La relation humain/chat était bénéfique pour le contrôle des rongeurs destructeurs de récoltes, qui avaient aussi uni leur destin à la civilisation humaine. Bien que de nombreux chats aient fini par devenir animaux de compagnie, le chat moderne n’est pas totalement domestiqué au sens classique du terme. Les chats modernes sont restés autosuffisants si c’est nécessaire, conservant de grandes compétences en chasse même quand on leur fournit leur nourriture et montrant un spectre de comportements allant d’animaux de compagnie indomesticables à extrêmement affectueux.
Les chats se sont répandus dans virtuellement toutes les parties du vieux Monde, probablement le long des routes marchandes qui reliaient les civilisations anciennes. Malgré leur expansion rapide, les chats sont restés étonnamment similaires à leurs ancêtres félins sauvages (les sous-espèces de Felis silvestris) dans leur aspect et leur activité. Les espèces souches des chats domestiques sont restées compatibles avec l’agriculture humaine. Le flux de gènes entre les chats sauvages et les chats domestiques modernes, et entre les chats domestiques modernes et leurs sous-espèces sauvages n’a pas eu d’impact négatif sur le rôle des chats comme principal petit carnivore dans les écosystèmes dominés par l’homme. En fait, avoir un pool sauvage de chats modernes autour des villages et des fermes peut avoir été bénéfique pour le contrôle des nuisibles et des zoonoses Une zoonose (du grec zôon,animal, et nosos,maladie) est une maladie animale transmissible à l' homme et vice versa. associées. De ce fait, l'envie de modifier les chats pour les faire correspondre à certains besoins des humains était certainement moindre que pour les autres espèces domestiquées et le développement des races de chats est resté considérablement à la traîne. Bien que les raisons de modifier l’aspect de base et la fonction du chat n’aient pas été aussi convaincantes que pour d’autres espèces, certains types de chats ont été sélectionnés artificiellement dans diverses régions du monde. Il est intéressant de noter que cette «sélection de race» était souvent fondée sur l’esthétique et concernait de simples caractéristiques de couleur de robe, de motifs colorés, etc. et moins visiblement la forme et la fonction. Les chats ne se sont pas prêtés à devenir bergers, employés à la chasse ou gardiens, mais leur grâce et leur beauté ont toujours été évidentes.
Le petit sous-groupe de chats domestiques qui ont subi une sélection artificielle intensive est celui des chats à pedigree (de pure race), qui ont été élevés pour conserver ou modifier des caractères purement esthétiques. Des 41 races reconnues par la Cat Fanciers’ Association (CFA), on pense que 16 «races naturelles» sont des variantes régionales qui précèdent la cat fancy. Les autres races ont été développées au cours des 50 dernières années et sont habituellement définies comme de simples variants génétiques dérivés des races naturelles. Cela contraste fortement avec la plupart des autres espèces domestiques qui ont subi des millénaires de sélection intense pour obtenir des caractères comportementaux, de performance ou de production qui mettent en jeu de complexes interactions de gènes.
Une étude phylogénétique récente conclut que le chat domestique est le produit d’un seul événement de domestication au Proche Orient ; cependant les modèles de dissémination du chat depuis cette domestication initiale n’ont pas encore été étudiés. Les objectifs du présent document sont triples. Le premier est de retracer le trajet du chat moderne à travers l’ancien monde et vers les Amériques, en confirmant ainsi les origines de la domestication. Le second objectif est de mesurer tous les changements de la diversité génétique qui ont peu se produire durant la migration des chats à partir du Croissant Fertile. Le troisième objectif est de mesurer toute perte de diversité génétique qui pourrait être imputée au développement de races anciennes (dites de fondation) ou plus contemporaines. Les données génétiques reportées ci-dessous proviennent de plus de 1100 chats, représentant 17 populations aléatoires dispersées dans cinq régions (Europe, Bassin Méditerranéen, Asie, Afrique, Amériques) et 22 races reconnues, comprenant des races de fondation et des races modernes (tableau 1). Des marqueurs microsatellites très polymorphes [3.] ont été utilisés pour déterminer les relations génétiques entre les races de chats, les origines géographiques et les niveaux de perte génétique dus à l’inbreeding (consanguinité). [7.]
Résultats
Phylogéographie
Elle étudie les phénomènes génétiques et démographiques, en particulier les phénomènes de spéciation qui ont conduit à la distribution et à la structuration actuelle des populations d'une espèce.Structure phylogéographique des chats domestiques
Une analyse bayésienne des populations spontanées, réalisée avec le logiciel STRUCTURE [4.], a élucidé la phylogéographie du chat domestique.
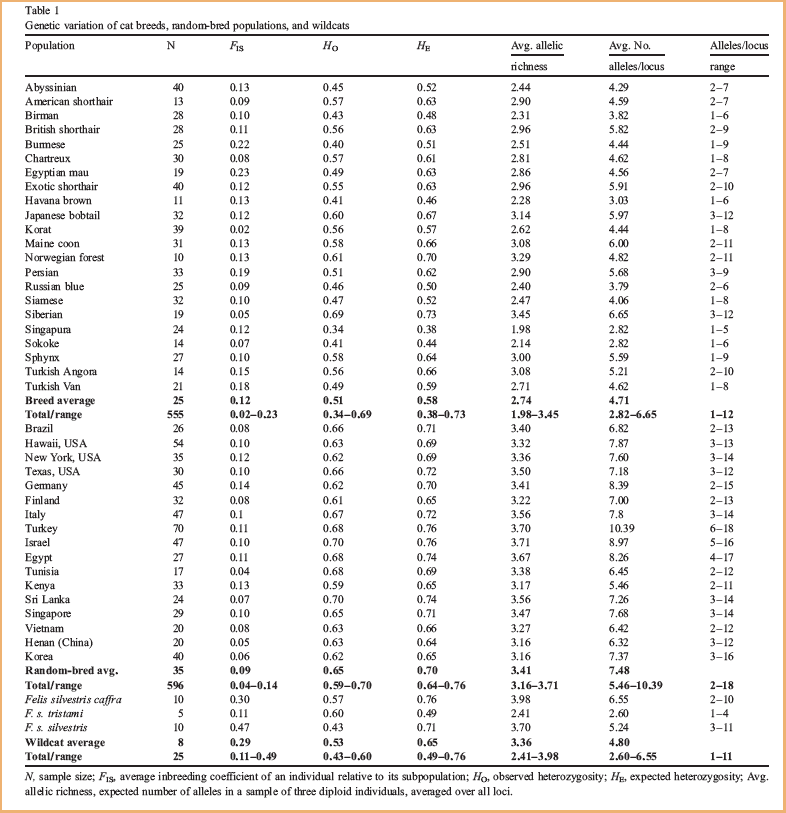
Les modèles de regroupement ont été examinés avec les groupes ancestraux (K) établis entre 2 et 4. La première scission (K=2) a séparé les chats d’Asie et d’Afrique de l’Est de toutes les autres populations (fig.1a).
Les populations du bassin méditerranéen ont été séparées des populations européennes à K=3. Les chats d’Afrique de l’Est ont formé un groupe [ndt : cluster] séparé et distinct à K=4. Le résultat de l’analyse de K=4 a clairement délimité quatre groupes génétiques de chats, correspondant à l’Europe, le Bassin Méditerranéen, l’Afrique de l’Est et l’Asie.
Figure 1b
(Haut) Première séparation de base de toutes les populations (K=2).Les races asiatiques, le Sokoké et les populations spontanées asiatiques (en vert) se distinguent clairement de tous les autres chats.(Bas) Analyse de toutes les populations (K=3).Les chats africains, méditerranéens et asiatiques (en bleu) se séparent des chats du sud-est asiatique (en vert) et des populations européennes (en rouge).
Nous avons ensuite intégré les races de chats aux analyses bayésiennes pour déterminer les relations les plus basiques entre toutes les races de chats et toutes les populations spontanées. A k=2, 4 des 5 groupes de chats spontanés d’Asie se sont reliés à 6 des 7 races asiatiques avec des probabilités de 75-85% ; ce regroupement de 10 populations est aussi distinct de toutes les autres populations de chats (fig.1b).
A K=3, les regroupements ont été affinés en groupe du Sud-Est Asiatique, groupe d’Europe de l’Ouest, et un groupe qui inclue tous les autres chats du Bassin méditerranéen, de l’Afrique de l’Est et de l’Asie du Nord. Les chats de Singapour sont un mélange de chats d’origines asiatiques et européennes. Les chats de gouttière américains se groupent avec ceux d’Europe de l’Ouest. Les chats italiens se sont révélés comme un mélange de fonds européen et méditerranéen. Les races du Sud-Est asiatique s'amalgament avec les populations sauvages du Vietnam. Parmi les races asiatiques, seul le Bobtail Japonais se rapproche fortement des chats occidentaux, bien qu’avec une influence asiatique ; Les deux races originaires des Etats-Unis (Maine Coon et American Shorthair) se rapprochent des sept races d’Europe occidentale. Etonamment le Persan est lié aux chats d’Europe occidentale. Le Sokoke, une race indigène africaine, est étroitement apparenté aux chats de gouttière du Kenya.
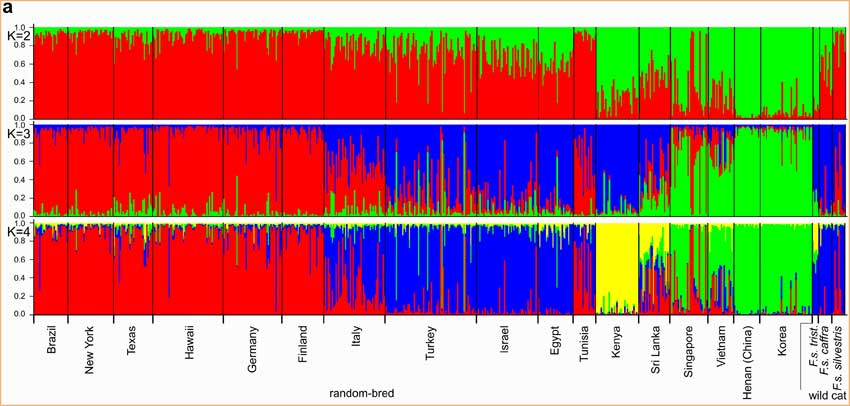
fig.1a Les 3 premières séparations de base des chats spontanés et sauvages
(Cliquez sur l'image pour l'agrandir)
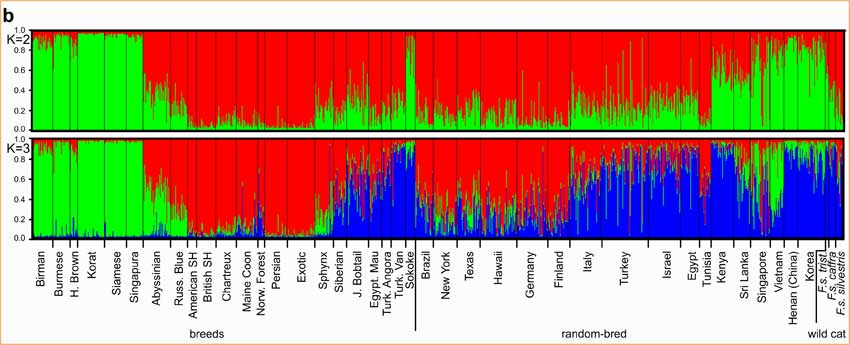
fig.1b Séparation (K=2) et analyse (K=3) de toutes les populations
(Cliquez sur l'image pour l'agrandir)
Figure 1c
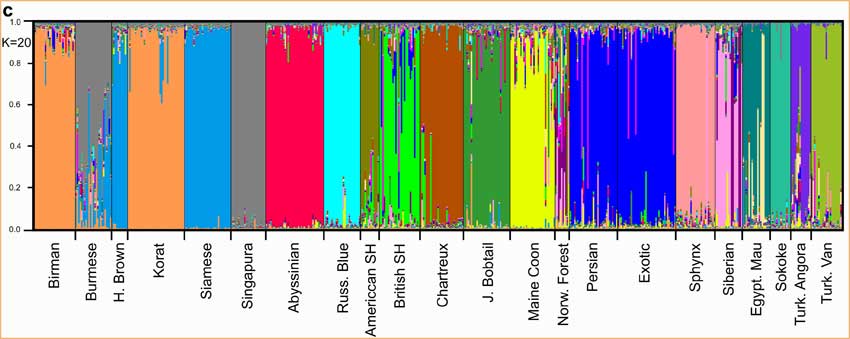
Trois paires, Havana browns et Siamois, Persans et Exotics et Burmese et Singapura ne sont pas différenciées génétiquement.Le Burmese semble partager des origines avec les Siamois et les Korats.
Les Korats et les Birmans apparaissent aussi comme étroitement liés.
Persans et Sibériens montrent la plus grande hétérogénéité intra-race.
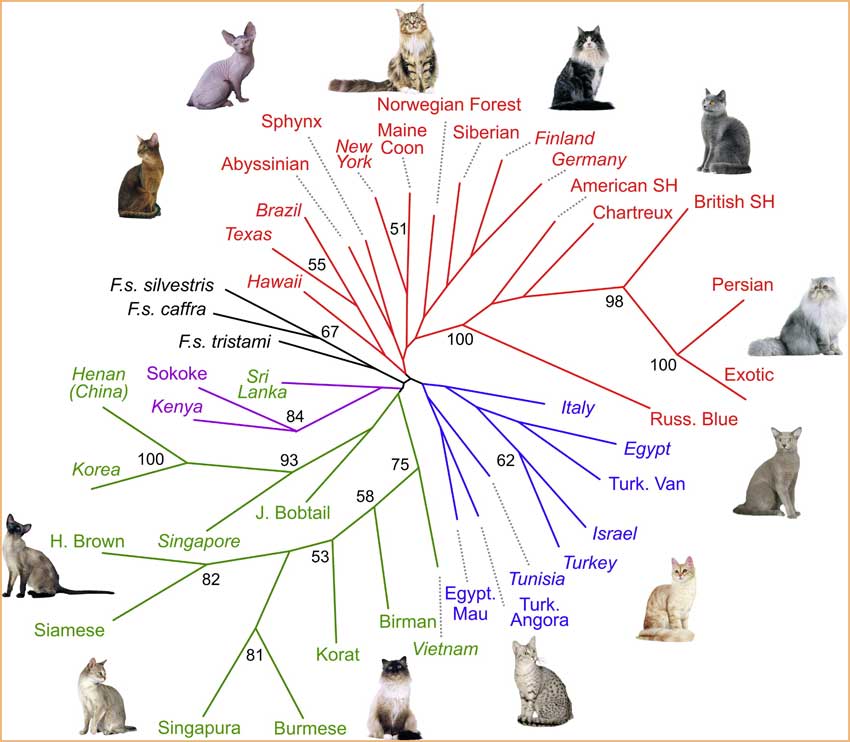
Nous avons construit des arbres phylogénétiques selon la méthode «neighbor joining» pour obtenir une confirmation supplémentaire de la structure génétique et comme autre moyen de représentation des modèles phylogéographiques.
L'utilisation de La mesure harmonique de Cavalli-Sforza et de la distance génétique de Nei ont donné des résultats similaires. [5.]
L'arbre produit par la mesure harmonique (fig.2) a un support plus fort, comme indiqué par les valeurs amorces supérieures à 50%, et différenciait le groupe asiatique de tous les autres chats. En outre, les groupes d'Afrique de l'Est, de Méditerranée et d'Europe forment chacun un embranchement monophylétique. Cette structure phylogéographique est aussi étayée par l'analyse factorielle des correspondances (FCA) (fig.3 et tableau 1).
Les chats de la périphérie de la mer d'Arabie, de Sri Lanka et les Abyssins relient le groupe européen et l'asiatique. Alors que les populations asiatiques sont très dispersées, les chats d'Europe occidentale forment un groupe serré.
Codage couleur des populations
Asie : vertEurope occidentale : rouge
Afrique de l'Est : pourpre
Bassin méditerranéen : bleu
espèces sauvages : noir.
Diversité génétique chez le chat domestique
La différenciation génétique a été déterminée par l'analyse de variabilité moléculaire (AMOVA) [6.] pour les populations de chats d'Europe (y compris les Amériques), d'Afrique de l'Est, de Méditerranée et d'Asie orientale (tableau 1). Les échantillons américains ont été inclus aux populations européennes, car aucun chat domestique n'est indigène des Amériques. La plus grande diversité génétique a été trouvée chez les chats de la région méditerranéenne (500 allèles parmi tous les marqueurs). Les chats méditerranéens ont aussi 41 allèles spécifiques, alors que les chats asiatiques en possèdent 14 (tableau 2).
Structure génétique des races de chats
Le regroupement bayésien [4.] a été utilisé pour examiner les relations génétiques entre les races. Une gamme de nombres de groupes a été étudié, et 20 a été choisi comme étant le plus approprié [NDT : K = 20]. Cette méthode a séparé les chats en groupes correspondant à leur race désignée (fig.1c) et avec une précision supposée de 95%. Les chats de 14 races ont été assignés à des groupes distincts par cette analyse statistique. La séparation de 8 races en 4 paires n'était pas cohérente. Le Singapura et le Burmese, le Havana Brown et le Siamois, le Korat et le Birman, et l'Exotic Shorthair et le Persan n'ont pas pu être identifiés comme des races distinctes. Cependant, ces 4 paires de races peuvent être différenciées sans équivoque quand on les analyse de manière indépendante des autres races (données non publiées ici). Deux races liées à la géographie (avec une origine géographique précise), l'Angora Turc et le Turc de Van sont distinctes l'une de l'autre. Cinq races (British Shorthair, Exotic Shorthair, Chat des Forêts Norvégiennes, Persan et Sibérien) présentent des subdivisions à l'intérieur de chacune d'elle, indiquant des origines multiples.
Modifications de la diversité génétique avec les races pures
Les races de chats ont une diversité génétique globale inférieure à celle des populations spontanées : la relativement faible composante de variance intra-population (Tableau 2) le met en évidence. L'hétérozygotie moyenne pour les animaux d'origine aléatoire et de pure race est respectivement de 0,65+/-0,03 pour les premiers et 0,51 +/- 0,09 pour les seconds. Des races telles que le Burmese, le Havana Brown, le Singapura et le Sokoke ont les valeurs d'hétérozygotie les plus faibles alors que le Sibérien a la plus élevée parmi les races, comparable à celle des populations spontanées. Le nombre moyen d'allèles par locus, le cœfficient de consanguinité moyen (Fis), et la richesse allélique suivent une tendance similaire (tableau 1, fig.4).
Argumentaire
Génétique de la diaspora féline
Il y a peu, de forts indices ont suggéré que la domestication du chat s'est produite dans la partie proche orientale du Croissent Fertile, suite au développement de l'agriculture. Le bassin méditerranéen avait été précédemment suggéré comme le site de la domestication du chat sur la base de preuves archéologiques. Driscoll et ses collègues ont utilisé une séquence mitochondriale et des marqueurs microsatellites pour élucider l'origine de la domestication du chat. Parce que les marqueurs microsatellites sont un meilleur indicateur d'une diversité génétique plus récente, cette étude s'est concentrée sur les marqueurs microsatellites pour évaluer les origines les plus récentes des races. Il ressort de la présente étude que la diversité génétique n'a pas été notablement diminuée par la dispersion ultérieure des chats modernes de la Méditerranée vers les autres régions de l'ancien monde. Cependant, il y a des différences régionales intéressantes dans leur constitution génétique. L'AFC, le regroupement bayésien et les arbres phylogénétiques neighbor joining ont divisé tous les chats du monde en quatre groupes distincts : Asie, bassin méditerranéen, Europe occidentale et Afrique de l'Est. Les chats américains se sont systématiquement regroupés avec ceux d'Europe occidentale, suggérant que les pionniers européens ont vraisemblablement introduit des chats dans le Nouveau Monde et que le temps de présence du chat en Amérique a été trop bref pour créer une différenciation génétique significative.
La diversité génétique est restée assez uniforme dans les diverses régions du pourtour méditerranéen. Le mouvement constant des bateaux et des caravanes, comme des chats, dans cette région clef de civilisation précoce, semble avoir favorisé un échange constant de chats. Bien que les chats méditerranéens tendent à être génétiquement uniformes, il y a quelques différences et des relations intéressantes entre certaines parties de la région méditerranéenne. L'analyse bayésienne indique que les chats italiens et tunisiens sont un mélange de chats d'Europe occidentale et de Méditerranée. Ce mélange est étayé par les liens historiques entre la Tunisie et les pays d'Europe Occidentale. Les chats de Sri Lanka et Singapour sont un mélange de chats du Sud Est Asiatique, d'Europe et de partout ailleurs quand ils sont définis par K=3 (fig.1b). Cependant, certains chats de Singapour n'apparaissent pas être des hybrides, mais plutôt un échantillonnage varié de chats génétiquement différents. Cela peut être un reliquat du colonialisme ou une importation récente. Selon l'AFC (fig.3), les chats sri-lankais et les Abyssins relient les chats de l'orient et de l'occident, peut-être à cause des voies commerciales maritimes de la mer d'Arabie ou encore du récent colonialisme britannique. La différence la plus intéressante concerne le groupe de chat asiatiques, qui est génétiquement distinct des chats du Bassin Méditerranéen, d'Europe occidentale et d'Afrique. Ce modèle de diversité génétique indique que les premiers chats domestiques ont atteint l'Extrême Orient, et que cela a été suivi par une longue période d'isolement relatif. Cet isolement peut avoir été causé par la croissance et le déclin du commerce entre les successions des grands empires anciens. La population asiatique présente un autre intérêt car elle a été divisée. Les populations de chats des différentes parties de l'Asie (spontanés et de race pure) sont génétiquement plus différentes l'une de l'autre que les populations locales des groupes du Bassin Méditerranéen ou d'Europe occidentale.
Figure 3
Les carrés représentent les centres de populations des régions géographiques : Asie (vert), Europe occidentale (rouge), Bassin méditerranéen (bleu), Afrique de l'Est (pourpre) et chats sauvages (noir).Les affinités des Sokokés avec les populations asiatiques, des Bobtails japonais avec les chats méditerranéens et européens, et des chats tunisiens avec les populations européennes sont évidentes, en accord avec les résultats de STRUCTURE.
Les espèces sauvages apparaissent comme plus distinctes des autres groupes.
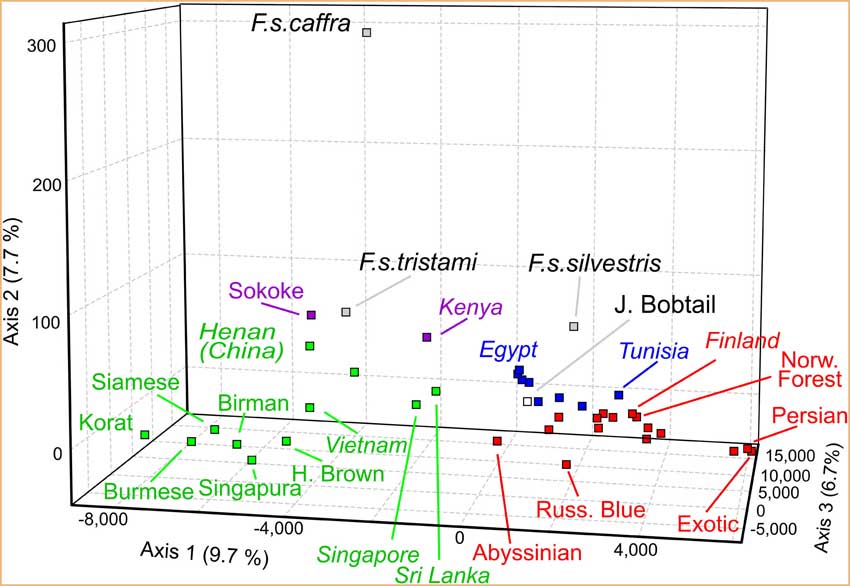
fig.3 Analyse de correspondance factorielle des races et populations
(Cliquez sur l'image pour l'agrandir)
Le cloisonnement de la diversité génétique au sein de la population asiatique est aussi corroboré par les longueurs des branches supérieures à l'intérieur de l'embranchement asiatique sur l'arbre phylogénétique (fig.2). Ceci suggère non seulement que la population asiatique a été relativement isolée des trois autres groupes régionaux, mais aussi que les chats des différentes régions d'Asie sont restés séparés.
La variation génétique dans les populations aléatoires et de pure race
L'infrastructure des populations est importante pour les analyses de parenté, les applications medico-légales, les appariements, et l' étude des maladies. [7.] Les populations spontanées présentent la plus grande variance intra-population, cohérente avec la taille plus importante de leurs populations, la liberté de migrer, et aucune sélection artificielle. Cependant, les différentes races de chats illustrent la variance inter-population la plus importante, reflétant leur diversité. La composante de variance intra-population des chats de gouttière est de 86%, comparé à 93-95% chez l'humain, suggérant que les populations de chats particulières sont génétiquement moins variables que les populations humaines. Les chats de race, avec une variance intra-population de seulement 61% ont une variabilité génétique inférieure à celle des populations aléatoires, ce qui est aussi reflété par les estimations d'hétérozygotie. L'hétérozygotie moyenne des races était de 10% inférieure à celle des populations aléatoires (fig.4) qui est elle-même inférieure de 10% à la moyenne des populations humaines. Les populations humaines du monde entier présentent une hétérozygotie moyenne de 0,50 à 0,78, la plupart étant au-dessus de 0,70. La variance intra-population intra-régionale des populations félines est environ le double de celle constatée chez les humains (6% chez les chats, 2,5% chez les humains), tout comme la variance intra-régionale (8% chez les chats, 3,6 à 4,3% chez les humains. Cette augmentation suggère que les populations félines sont plus isolées génétiquement à cause de la géographie que les populations humaines. L'importante variance intra-population (24%) et intra-régionale (15%) des races est vraisemblablement le résultat de l'intense sélection artificielle et de l'isolement imposés par les pratiques d'élevage.
| TABLEAU 2 | |||
| AMOVA des races et des populations spontanées | |||
| Composants de variabilité | |||
| Echantillon | Entre les populations | Dans les populations /entre les régions | Dans les régions |
| Races | 61 | 24 | 15 |
| Populations spontanées | 86 | 6 | 8 |
| Données combinées | 73 | 18 | 9 |
Les races Burmese et Singapura ont l'hétérozygotie la plus basse et le plus grand Fis [ndt : cœfficient de consanguinité] de toutes les races, reflétant l'inbreeding le plus intensif (fig.4). La race la plus récente de la CFA, le Sibérien, a la variation la plus élevée, comparable à celle des populations aléatoires. Cela indique qu'elle est assise sur une grande quantité de chats de fondation. Le Sphynx, qui dérive du Devon Rex, au aussi une forte diversité génétique. Au vu de ces résultats, les éleveurs de Burmese et de Singapura devraient prêter attention à la diversité génétique, alors que les éleveurs de Sibériens devraient être encouragés à conserver la diversité existante pendant que leur race se développe. Comme pour les efforts de conservation des populations de félins exotiques captifs, les analyses génétiques, l'histoire des races et la dynamique des populations pourraient être utilisées pour développer la gestion de la race ou des programmes de survie pour conserver la variation génétique au sein du pool génétique (sur le long terme) pour une période de longue durée. En outre, ces différences régionales dans la variabilité génétique des chats suggèrent que les marqueurs génétiques utilisés pour l'établissement du profil des chats devraient être testés dans les diverses populations pour valider leur efficacité.
Figure 4
Les barres claires représentent l'hétérozygotie observée (HO) et les barres noires le cœfficient d'inbreeding (FIS).Les populations sont classées suivant l'ordre croissant d'hétérozygotie, les races à gauche, les populations spontanées au milieu et les espèces sauvages à droite/
L'axe des y représente la proportion de l'hétérozygotie ou du cœfficient d'inbreeding.
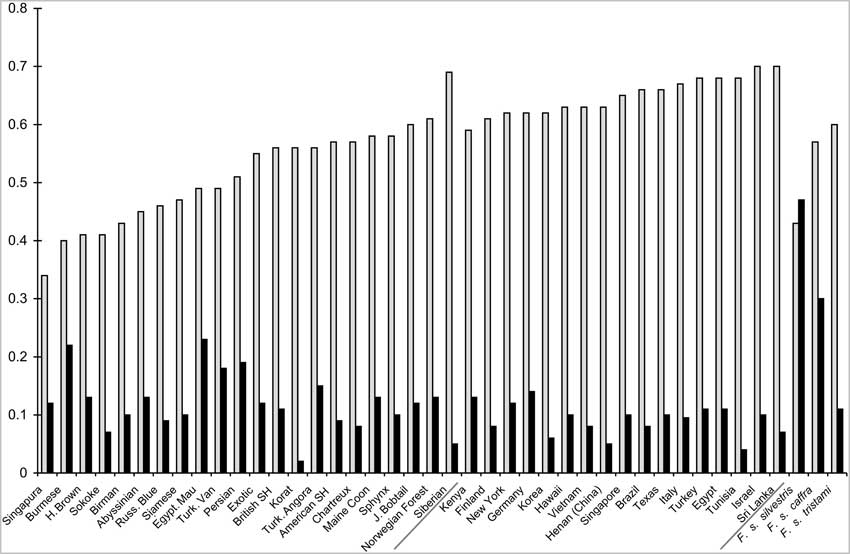
fig.4 Indices de diversité génétique des races, populations spontanées et chats sauvages.
(Cliquez sur l'image pour l'agrandir)
Structure génétique et origine des races de chats
Nous avons démontré que la diversité génétique qui a suivi la domestication était localisée dans les régions de l'ancien monde et même dans des localités à l'intérieur de ces régions. Certaines différences peuvent être expliquées par l'isolement, alors que d'autres se comprennent mieux par les échanges d'animaux entre les groupes et les localités. Toutes ces différences impliquent des interventions humaines, mais elles n'étaient probablement pas intentionnelles. Cependant, les modifications volontaires de la génétique des chats se sont finalement produites au cours du développement des races.
Contrairement aux races de chevaux domestiques, de chiens, de bovins, d'ovins etc., dont certaines ont des milliers d'années, la plupart des races de chats ont été développées au cours des 50 dernières années, essentiellement en Europe et aux Etats-Unis. Le Persan, le Bleu Russe, le Siamois et l'Angora font partie des premières races de chats enregistrées par les associations félines. La CFA désigne 16 races comme «naturelles» ou «de fondation». Quinze des races de fondation sont incluses dans cette étude, il y manque seulement le Manx, la race de chat sans queue qui s'est développée sur l'Ile de Man.
Arbre phylogénétique
Le vocabulaire fait référence à… Un arbre !Les nœuds représentent les ancêtres communs (inconnus) qui ont transmis à tous leurs descendants leurs caractères.
Les branches représentent les lignées issues de ces ancêtres qui se sont différenciées par l'acquisition de caractères différents. Leur longueur, quand l'arbre est construit d'après des distances moléculaires, traduit le temps écoulé en relation avec les mutations accumulées.
Et chaque branche se termine par une feuille, un groupe ou une espèce.
Les chats de pure race ont une structure similaire à celle des chats des populations spontanées de leur région. Les races du Sud-Est Asiatique, y compris le Birman, le Burmese, le Havana Brown, le Korat, le Siamois et le Singapura forment un regroupement distinct et à l'extrémité opposée du spectre génétique des races occidentales, telles que décrites par la CFA. Les Abyssins et les Bobtails japonais sont des exceptions et possèdent des marqueurs génétiques communs aux races asiatiques et occidentales. Cela indique que des chats d'Asie et d'Europe ont été utilisés pour créer ces races.
Chaque race de fondation est génétiquement distincte des autres avec 95% d'exactitude. L'une des races de chats les plus anciennement reconnues, le Persan, a été utilisée pour le développement de plusieurs autres races. Le croisement avec les Persans est souvent utilisé pour produire un type de tête plus brachycéphale. L'Exotic Shorthair est essentiellement une version à poil court du Persan, ce que prouve le regroupement complet de ces deux races en un seul groupe. La race la plus dolichocéphale, le Siamois, a aussi été utilisée pour créer de nombreuses autres races, notamment les Colourpoints, les Orientaux, les Havana Brown et leurs variantes à poil long. Bien que listées comme différentes, la plupart de ces races dérivées ne diffèrent que par une variante d'un gène unique, comme la longueur du poil, les motifs ou la couleur de la fourrure. En outre, ces races dérivées sont vraisemblablement impossible à isoler comme des races distinctes, comme cela a été montré pour le Siamois et le Havana Brown. Le regroupement du Burmese et du Singapura sont un exemple supplémentaire de récentes dérivations de races. L'étroite association du Persan et de l'Exotic, du Siamois et du Havana Brown, du Burmese et du Singapura se concrétisent par les branches longues depuis le nœud dans l'arbre neighbor joining et l'impossibilité de séparer les membres de ces paires par les marqueurs génétiques utilisés dans cette étude.
Cependant, il est important de se rappeler que la plupart des races ont une histoire orale/écrite outre leur empreinte génétique. Le folklore de l'élevage du chat prétend que le Burmese et le Singapura sont indigènes à l'ancienne Birmanie. Le folklore suggère aussi que des Burmese des Etats-Unis ont été amenés à Singapour, croisés volontairement avec des chats indigènes, et ensuite ramenés aux Etaits Unis en tant que nouvelle race Singapura. Les Havana Brown sont considérés comme une race à part aux Etats-Unis, cependant les associations félines européennes les considèrent uniquement comme une variante de couleur des Siamois.
Ces données montrent que la plupart, mais pas toutes les races de fondation ou modernes, semblent être issues des populations spontanées de leur supposée région d'origine. Les races du Sud-Est Asiatique sont fortement associées aux populations spontanées du Vietnam, de Chine, de Corée et de Singapour. Le Sibérien est une des races les plus récentes de Russie et est activement travaillée. Nous n'avons pas pu avoir de chats des populations spontanées de Russie, mais le Sibérien est très fortement associé aux chats des pays voisins d'Allemagne et de Finlande. Les Turcs de Van se regroupent avec les chats des populations aléatoires régionales de Turquie, Israël et Egypte. De même, les chats des Iles du Kenya et Lamu et Pate se lient avec la race Sokoke, qui a été définie au Kenya. On accédait aux îles kenyanes et au Kenya continental par les routes commerciales de la mer d'Arabie, ce qui permettait les échanges entre l'Inde, le Proche-Orient, la Péninsule Arabique et l'Afrique. Toutes les autres races et les populations spontanées forment un réseau de chats fortement apparentés et moins individualisés du groupe d'Europe occidentale. Etonnamment, le Persan n'est pas associé génétiquement avec les populations spontanées du Proche-Orient, mais se joint à celles d'Europe occidentale. Le Persan est peut-être la plus ancienne race de chat connue, et a subi une sélection en vue de l'obtention d'un phénotype extrême qui a probablement impliqué des interactions de gènes complexes. Bien que le Persan des débuts puisse être réellement originaire de l'ancienne Perse, le Persan moderne a perdu sa signature phylogéographique. De même, le Bobtail japonais ne se révèle pas être issu de ses origines déclarées du Japon. Bien que les chats ne soient pas indigènes au Japon, ils ont migré vers ces îles qui faisaient partie des routes commerciales d'Asie il y a des centaines d'années. Introduit comme race aux Etats-Unis en 1968, le Bobtail japonais des Etats-Unis présente un patrimoine génétique plus influencé par les chats européens que par les chats asiatiques. Le Mau Egyptien est également près de perdre ses origines historiques via des influences génétiques européennes. Globalement, ces résultats indiquent que les populations spontanées et les races se regroupent d'une manière qui concorde largement avec la géographie. L'histoire des races est exacte pour sa plus grande partie.
Cette étude a reconfirmé que le Bassin Méditerranéen est le site probable de domestication du chat moderne. On a trouvé des groupes de chats génétiquement distincts dans le Bassin Méditerranéen, en Europe, en Asie, et en Afrique. Les chats d'Amérique du Nord sont étroitement apparentés aux chats européens. Le groupe asiatique est génétiquement unique, indiquant qu'il est devenu isolé après son introduction. La diversité génétique est restée étonnamment large parmi les chats de diverses parties du monde. Les données génétiques prouvent que la plupart des races de fondation, mais pas toutes, sont issues des chats indigènes de la région d'origine supposée et que leur naissance est associée à une perte de diversité. Les chercheurs doivent reconnaître l'aspect récent du développement des races, qui agit sur le «Linkage disequilibrium» [NDT : similitude des allèles de microsatellites situés au même locus. Se mesure en fréquence dans les populations. Une grande fréquence peut être associée, en raccourci, à une moindre diversité allélique qui rejoint la notion de consanguinité, avec ses risques dans l'expression de certaines pathologies] et la grande utilité des études de croisement [NDT : ce qui suggère de maintenir la diversité allélique].
Cette étude est également un avertissement pour les éleveurs de chats actuels : le développement d'une race doit être fait lentement en conservant une large base génétique. On a identifié plus de 20 désordres génétiques néfastes chez les chats modernes, et tous ont été identifiés chez les races pures. Les relations génétiques identifiées entre les races de chats peuvent être utilisées par les éleveurs pour élaborer des programmes d'élevage des races plus efficaces.
Matériels et méthodes
[1.]
Les microsatellites sont de courtes séquences de l'ADN non codant comparées ici entre les différentes races et populations félines étudiées. Le nombre de différences repérées représente un nombre de mutations traduit en distances génétiques. Plus la distance est grande, plus le temps depuis lequel 2 races ont divergé est grand, temps proportionnel au nombre de mutations qui se sont produites.
Une page illustrée sur les microsatellites.
[2.] Collection d'échantillons et prélèvement d'ADN.
Notre base de données comporte 22 races de chats, incluant 15 à 16 races supposées de fondation, 2 races en développement, Siberien et Sokoke, et 5 races plus récemment crées, incluant Exotic Shorthair, Havana Brown, Bobtail Japonais, Singapura, et Sphynx (Table 1). Nous avons obtenu les échantillons d'ADN de la plupart des races lors d'expositions félines et par demande auprès de propriétaires de chats aux Etats Unis. Les données pour le Korat, l'Angora Turc, le Turc du lac de Van et le Sibérien proviennent des Etats Unis et d'Europe car ces races ont le même standard dans ces continents. Les pedigrees ont été vérifiés afin de s'assurer qu'il n'y avait pas de parentés de premier degré incluses dans l'étude et qu'une majorité de chats n'avaient pas de grands parents en commun. Les populations spontanées incluaient des chats sauvages n'ayant ni nourriture ni hébergement donnés par les humains, des populations ayant quelques contacts humains et des chats vivant avec des humains mais sans race précise. Pour les populations spontanées, nous n'avons pas retenu des portées ou des parents et leurs enfants…
… Les échantillons d'ADN du chat sauvage africain (F. s. caffra) et European (F. s. silvestris) sont issus d'études précédentes…
… L'ADN du chat sauvage d'Arabie (F. s. tristami) a été collecté en Isræl…
[3.] Génotypage Trente huit microsatellites ont été séquencés/comparés chez 1176 chats représentant 22 races, 17 populations spontanées et 3 populations sauvages. En moyenne, chaque population spontanée et de pure race était respectivement représentée par 35 et 25 chats, de 10 à 70 pour les chats domestiques. Pour le populations sauvages, 5 à 10 échantillons ont été séquencés. Les marqueurs microsatellites ont été choisis sur les critères de haute hétérozygotie, haut contenu informatif de polymorphisme et large distribution chromosomique…
[4.] Analyse structurale des populations
NDT : Cette annexe traite de l'exploitation des données. Du fait de leur très grand nombre, celle-ci est faite par des logiciels qui appliquent des algorithmes mathématiques. C'est assez «aride» ! Vous nous pardonnerez le raccourci que nous vous proposons.
La structure de toutes les populations échantillonnées a été déterminée par FCA (logiciel GENETIX 4.05 ) et par classification bayésienne [STRUCTURE (Pritchard et al., 2000), Algorithme permettant de classer N individus en K populations].
Les données analysées sont celles qui ont été recueillies par la comparaison des 38 microsatellites (donc de leurs allèles) dans les groupes choisis. Le logiciels analyse ces données sur la base de probabilités choisies à priori (suivant des données biologiques) et du nombre de groupes, K, dans lesquels on lui demande de les répartir. Par exemple :
- 1. La probabilité de mutation d’un nucléotide est uniforme.
- 2. Passer de A à G ou de C à T est plus probable que d’autres transitions.
- etc…
Le logiciel établit une probabilité «à posteriori», ici de regroupements de races en groupes, donc de parenté plus ou moins grande entre les différentes populations de chats étudiées, afin de retracer l'ancienneté de séparation de ces groupes. Le nombre de groupes K testé a été arrêté à 20… « Nous avons choisi à posteriori K=20 comme le meilleur indicateur car, pour cette valeur, les groupes génétiques formés par l'analyse correspondaient le plus aux races supposées»…
La filiation entre les groupes la plus probable permet de construire l'arbre phylogénétique le plus probable.
La méthode Bayésienne utilisée est celle gérée par le logiciel GeneClass2.
NDT : Là encore, une certaine aridité… Et donc une version simplifiée.
Les arbres phylogénétiques ont été construits à partir de la totalité des données. Elles ont été traitées par le programme PHYLIP (ou là) qui permet la construction d'arbres phylogénétiques suivant différentes méthodes. Ainsi, les termes de Nei ou Cavalli Sforza correspondent à deux indices de distance différents qui ont été développés dans le but de quantifier les divergences génétiques entre unités taxonomiques proches.
Pour les adeptes : Ont été employés dans l'ordre Seqboot, Genedist, Neighbor et Consense dont nous vous laissons le soin de découvrir les vertus grâce à quelques clics.
Dans chaque cas, les arbres obtenus ont été soumis à vérification : 1000 itérations ou répétitions d'un processus de vérification des résultats fournis par les algorythmes.
Le logiciel TreeView a été utilisé pour visualiser les arbres construits.
Pour quantifier le degré de différenciation régionale, une étude AMOVA a été conduite en 3 parties : Seulement les races, seulement les populations spontanées puis ces données combinées, à l'exclusion des chats sauvages.
Les populations ont été divisées en 4 groupes correspondant à leur origine géographique comme illustré par le tableau 3. L'origine géographique, à priori, des Abyssins et Chats de Sri Lanka, aussi bien avec les analyses par FCA et STRUCTURE, n'était pas claire. Aussi, ils n'ont pas été inclus dans l'étude AMOVA.
[7.] Paramètres des populations
Nous avons calculé les nombres et les variétés des allèles de la (des) population(s) de base et l'hétérozygotie observée et attendue (HO et HE respectivement) en utilisant GENALEX. Le cœfficient d'inbreeding (Fis) a été calculé en utilisant Arlequin. L'hétérozygotie attendue est celle dont on fait l'hypothèse suivant la loi de Hardy-Weinberg. La richesse allélique, mesure du nombre d'allèles standards pour une taille d'échantillon (3 individus), a aussi été calculée avec Fstat.
Les paramètres spécifiques des locus sont présentés dans le tableau 2.

Les routes du chat. La traduction d'une étude plus simple sur l'origine des populations de chats.