Cette étude présente un énorme intérêt dans le contexte actuel où il y a débat quant à la polydactylie du Maine Coon. Il y a d'une part les tenants du fait qu'elle se résume à la seule présence de doigts supplémentaires, sans autres conséquences négatives sur la morphologie ou la santé du chat. Il y a d'autre part ceux qui invoquent d'autres anomalies moins bénignes potentiellement associés à cette polydactylie.
Les uns acceptent donc d'élever des MC polydactyles et œuvrent à leur reconnaissance en exposition. Les autres tentent de s'opposer à cela.
L'étude conduite par l'équipe d'Edinburgh est relativement complexe. Pour cette raison j'ai fait le choix d'en faire un résumé assorti de commentaires qui me semblent permettre d'en faciliter la compréhension.
Ceci signifie en clair que j'ai pris beaucoup de libertés quant à la forme et l'ordre du contenu (dont j'ai volontairement écarté certaines parties). Ce travail peut donc être qualifié d'«interprétation personnelle». Cependant, liberté a été prise avec les mots tout en respectant totalement le contenu scientifique de cette étude et ses conclusions.
Mon commentaire strictement personnel figure à la suite de ce travail sous «Le mot de…»
Des mutations ponctuelles dans un cis-régulateur du gène sonic hedgehog génèrent une variation de régulation responsable de la polydactylie préaxiale
Introduction
[ndt : Elle pose le contexte de l'étude et définit son cadre génétique. En cela, les données qu'elle contient sont essentielles à la compréhension de ce qui suit, protocoles expérimentaux adoptés et interprétation des résultats obtenus, d'où certains renvois en bas de page.]
La construction du membre pendant l'embryogenèse, dont sa partie distale, main ou pied, fait intervenir en cascade de nombreux acteurs, gènes et leurs séquences régulatrices, donc de nombreux intermédiaires moléculaires codés par ces gènes ou séquences ADN. Elle est ainsi l'objet d'une régulation très fine qui conduit à la construction parfaite du membre suivant le plan propre à l'espèce.
Dans le génome des Vertébrés, une série de séquences non codantes, dites CNE sont remarquablement conservées [1.]. Ceci signifie qu'on les trouve dans le génome de toutes les espèces, avec très peu de modifications de leur séquence. Dans de nombreux cas, ces séquences sont des éléments régulateurs cis, ce qui veut dire qu'elles régulent l'activité des gènes [2.]. Le signal moléculaire sonic hedgehog ou Shh [3.] est ainsi régulé par un certain nombre de ces CNE qui contrôlent l'expression du gène correspondant Shh. Shh intervient notamment dans la construction du membre à partir de son bourgeon.
L'un de ces CNE, appelé ZRS [4.], est le régulateur cis qui contrôle l'expression de Shh lors du développement du membre. Le ZRS s'active spécifiquement dans une zone située dans la marge postérieure du bourgeon du membre et appelée ZPA. A travers la production de Shh, la ZPA régule l'identité et le nombre de doigts qui seront construits suivant l'axe antéro-postérieur de la main qu'elle définit : Pôle antérieur côté pouce et pôle postérieur côté petit doigt. [5.]
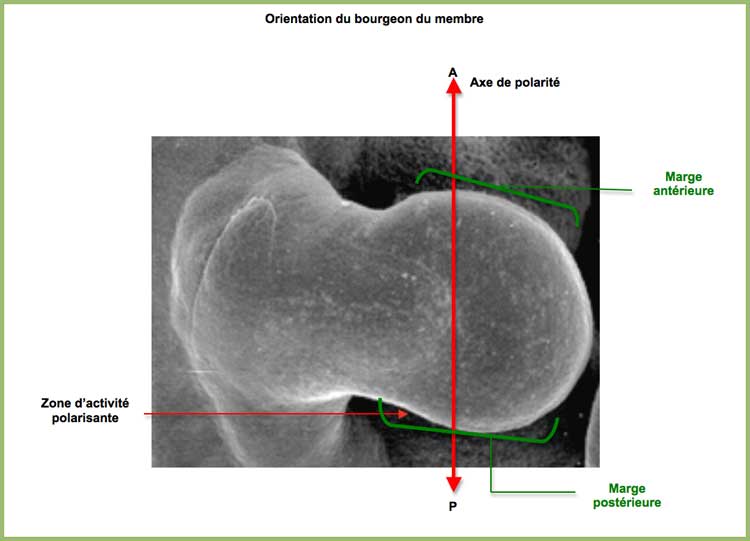
Polarité du bourgeon du membre et ZPA
(Cliquez sur l'image pour l'agrandir)
La place de la ZPA détermine l'axe de polarité du membre et donc de sa partie distale, main ou pied. L'expression des gènes construisant ce segment distal est ainsi orientée de façon à construire une main ou un pied non symétrique (doigts de tailles différentes).
Le rôle de la ZPA dans la construction des doigts est démontré par le résultat d'une greffe dans la marge antérieure du bourgeon du membre chez un embryon précoce : Il y a construction d'une main en miroir.
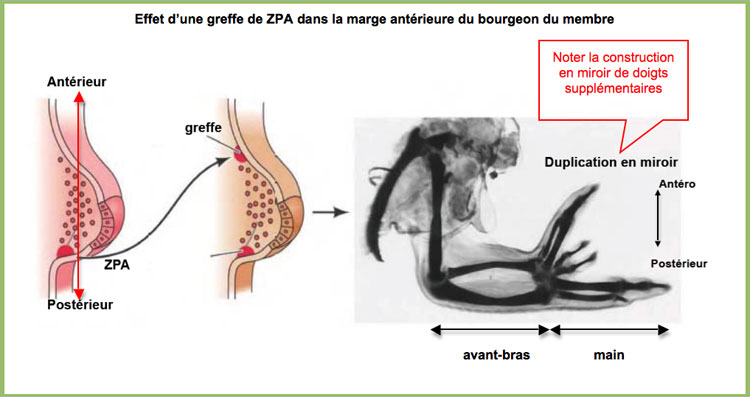
Effet d'une greffe de la ZPA
(Cliquez sur l'image pour l'agrandir)
Les données valident le fait que la polydactylie préaxiale, notamment chez l'Homme et les modèles Souris, résulte de mutations dans ZRS. Celles-ci aboutissent à une anomalie d'expression de Shh qui génère une ZPA ectopique
Littéralement «en dehors du lieu normal». avec, en conséquence, la formation de doigts surnuméraires.
L'étude de chats polydactyles, dont les célèbres chats d'Hemingway, ajoute de façon significative au nombre de mutations associées à cette anomalie.
ZRS, ce cis-régulateur de Shh, a la singularité de présenter des mutations qui modifient son activité et ainsi son expression spatiale [ndt : Le lieu de l'embryon où il s'exprime]. Avec les expériences réalisées sur des souris transgéniques, les chercheurs ont montré que les mutations donnent lieu à des expressions très variables. Elles dépendent de la position et de la nature de la mutation dans ZRS. Dans nombre de cas, l'expression modifiée par la mutation est observable dans le phénotype. Des mutations ponctuelles spécifiques conduisent à des changements de morphologie du membre qui tendent à valider la notion que de nouvelles formes morphologiques ne nécessitent pas des modifications moléculaires importantes. Une modeste modification de régulation peut transformer un caractère structural fondamental chez un Mammifère.
Résultats
Des mutations dans ZRS causent la polydactylie des chats domestiques
Listons les mutations étudiées dont le schéma vous donne la position relative et la nature dans la séquence CNE (ZRS) :
La séquence ZRS sauvage (non mutée, notée Wt) a été déterminée à partir de 5 chats non polydactyles. Les cinq n'ont montré aucune différence dans leurs 800 paires de bases.
ZRS a été séquencé dans l'ADN provenant de chats d'Hemingway [6.], colonie où est présente la polydactylie. Tous les polydactyles d'Hemingway portent la même mutation en position 479 : Substitution de l'adénine A par la guanine G. Elle a été appelée mutation Hw (forcément ! HemingWay !) et détermine majoritairement une polydactylie des pattes avant.
Le séquençage du ZRS chez des chats britanniques a permis de découvrir 2 autres mutations, UK1 et UK2 (United Kingdom&hellip De la simplicité des biologistes !). UK2 est d'ailleurs très proche de Hw : Elle affecte le même codon (triplet) mais un nucléotide différent. Ce codon est conservé alors que cette partie de la séquence présente par ailleurs une grande variabilité dans les génomes des Mammifères.La mutation Hw n'a pas été trouvée chez les chats britanniques.
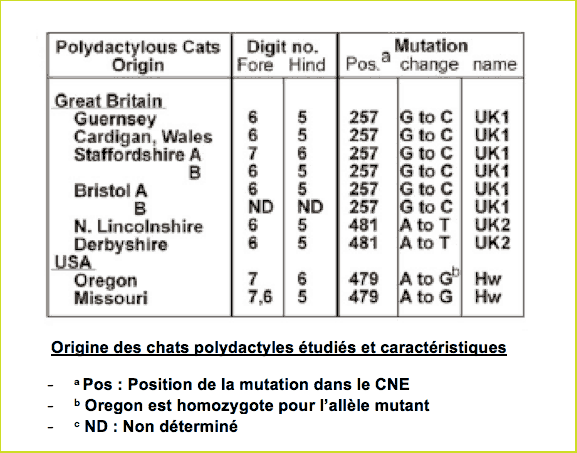
Origine et caractéristiques des chats étudiés
(Cliquez sur l'image pour l'agrandir)
D'autres mutations ont été découvertes dans ZRS de l'Homme et de la souris : Cuban, belgian et family B chez l'Homme, M 100081 chez la souris (forcément ! M pour Mouse!)
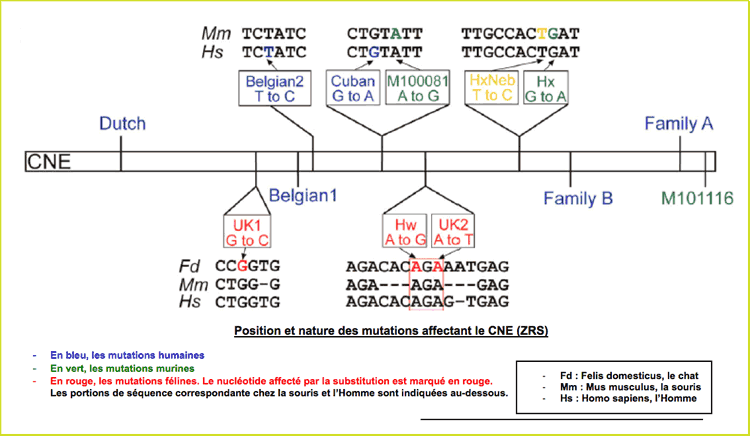
Effet d'une greffe de la ZPA
(Cliquez sur l'image pour l'agrandir)
Toutes sont des mutations ponctuelles qui affectent une seule paire de bases. Les phénotypes déterminés vont de la simple polydactylie, cas pour Hw, à des anomalies bien plus graves et étendues dans le cas de Cuban par exemple (et dont je vous passe le descriptif).
La question était de préciser comment ces mutations déterminent des éléments squelettiques additionnels (seulement des doigts ou d'autres anomalies squelettiques associées) et notamment cerner l'expression et les conséquences de la mutation Hw qui est celle isolée chez le MC.
Les protocoles expérimentaux
l'étude a été réalisée sur des souris de génotype sauvage et des souris transgéniques. Celle-ci ont reçu un transgène comprenant une séquence ZRS sauvage ou mutée afin d'en suivre l'expression. Pour repérer les zones et l'intensité avec laquelle ZRS s'exprime, un gène reporter qui est en quelque sorte un «mouchard» a été attaché à la séquence CNE. Quand le fonctionnement de ZRS active le reporter, ce dernier crée une réaction enzymatique qui transforme un substrat en lui donnant une couleur bleue. Dans le bourgeon de membre, le lieu d'activité de ZRS est donc repérable par une coloration bleue dont l'intensité traduit le degré de fonctionnement.
Résultats expérimentaux et premières conclusions
Une première expérience a consisté à étudier les conséquences des mutations de l'Homme et de la souris : Mutations Cuban et Belgian2 pour l'Homme, M 100081 et Hx pour la souris.
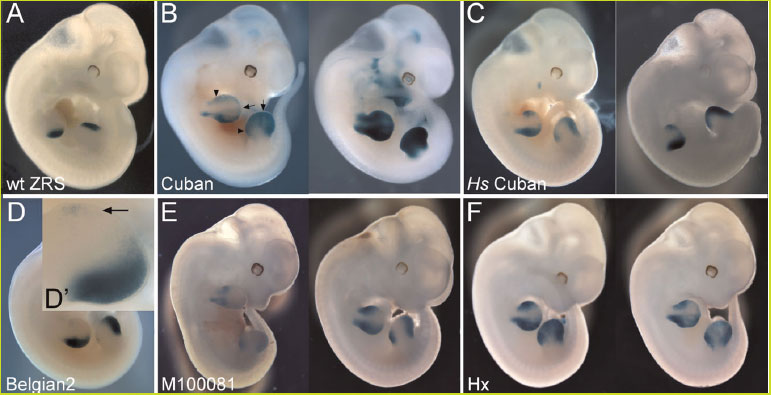
Analyse transgénique des mutations humaines et murines
(Cliquez sur l'image pour l'agrandir)
Par rapport à l'embryon non mutant, les embryons transgéniques montrent tous une activité du ZRS plus ou moins modifiée, en lieu et/ou intensité. L'embryon sauvage montre une activité localisée uniquement à la ZPA de la marge postérieure des bourgeons des pattes antérieures et postérieures. Belgian2 se traduit par une faible activité dans la marge antérieure. Elle est nettement plus importante pour la mutation Hs. Pour Cuban, on constate une activation dans les bourgeons des membres mais aussi à d'autres niveaux de l'embryon (dorsal notamment), ce qui explique les anomalies plus importantes affectant d'autres parties de l'organisme.
Cette étude montre déjà que l'expression de ZRS est la même chez l'Homme et la souris puisque les transgènes humains se sont exprimés dans les embryons de souris. Le phénotype humain résulte de l'expression ectopique de Shh pendant la formation des membres.
En second lieu, les expériences suggèrent qu'il y a une relation entre une mutation spécifique et le phénotype observé. Les différences constatées entre deux embryons porteurs de la même mutation illustrent la variabilité de l'expression selon les individus.
La seconde série d'observations a été faite sur deux lots de souris qui diffèrent par leur type de transgène. Celles notées Fd (pour Felis domesticus, le chat domestique) ont un transgène félin (ZRS de chat) sauvage, Hw ou UK2. Celles notées Mm (pour Mus musculus, la souris) ont un transgène murin, un ZRS de souris dans lequel a été placée une mutation féline à sa place exacte. Ce protocole teste donc le rôle joué par les caractéristiques d'espèce de ZRS du chat ou de la souris dans l'expression des mutations félines Hw et UK2.
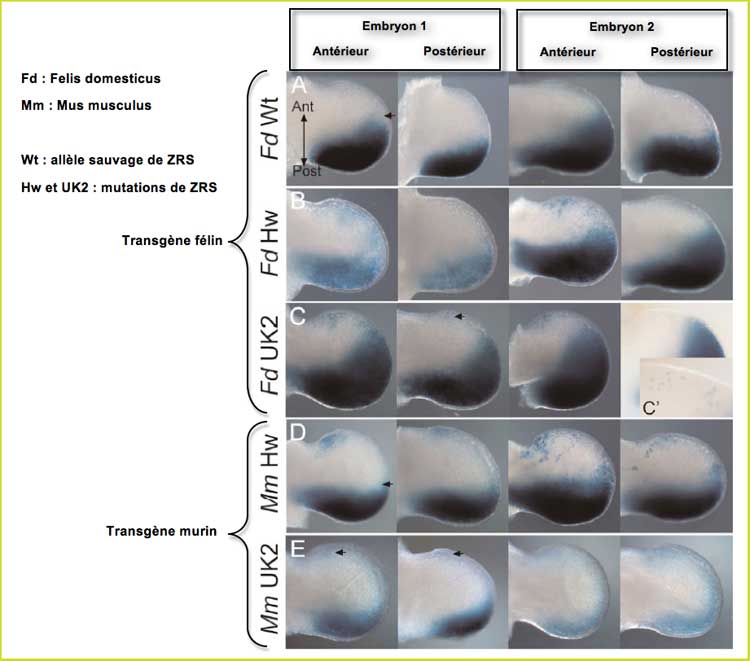
Analyse transgénique des mutations félines
(Cliquez sur l'image pour l'agrandir)
En B, on constate que Hw entraîne une activation antérieure du ZRS plus importante dans le bourgeon de patte avant que dans celui de patte arrière. Ceci est à relier au fait que chez les chats polydactyles d'Hemingway 80% le sont uniquement aux pattes avant.
En C, pour UK2, l'expression ectopique antérieure affecte les quatre membres. De fait, tous les polydactyles porteurs de UK2 le sont aux quatre pattes.
Quand la même mutation féline est contenue dans ZRS de souris, on constate un degré d'expression différent. En D, l'activation antérieure est plus importante dans le bourgeon de patte avant, et elle est aussi manifeste dans le bourgeon de patte postérieure. Pour UK2, en E, il y a une moindre activation dans le membre antérieur comme dans le postérieur.
Deux mutations ponctuelles très proches comme le sont Hw et UK2 conduisent à des expressions ectopiques différentes. Ces expressions sont corrélées aux phénotypes des membres (polydactylie avant ou avant + arrière).
L'expression d'une même mutation dépend aussi du contexte d'espèce du ZRS : Le triplet affecté est essentiel pour l'expression mais les différences peuvent être reliées au fait que certaines parties de la séquence ZRS ne sont pas conservées entre le chat et la souris.
L'ensemble de ces résultats illustre la plasticité des mécanismes de régulation qui interviennent dans la construction des membres, plasticité possible grâce à des différences de séquences, du fait des mutations ou des variations propres à l'espèce.
Discussion
L'étude des chats polydactyles a permis d'identifier trois nouvelles mutations associées à la forme préaxiale. Comme chez l'humain et la souris, ces mutations affectent le cis-régulateur ZRS, suggérant que la polydactylie chez le chat est équivalente à celle observée chez les autres Mammifères.
strong>Comme ces mutations produisent chez l'Homme un phénotype spécifique aux membres, sans autre anomalie physiologique discernable, nous proposons le fait que ce type de polydactylie n'a pas d'incidence nuisible sur la santé du chat.
La notion qu'une mutation ponctuelle puisse modifier un mode d'expression aussi bien que le fait l'inactivation d'un gène est une étape essentielle dans la compréhension de processus pathologiques. En considérant d'autres études génétiques de la région régulatrice de Shh, un peu de lumière est jetée sur le mécanisme d'activité du ZRS. La combinaison des données de l'étude transgénique et de l'étude de la délétion ciblée du ZRS [7.] démontre que le régulateur est nécessaire et suffisant pour déterminer le mode d'expression spatial de Shh dans le membre. De plus, les souris qui sont hétérozygotes pour la délétion ZRS ne présentent aucune anomalie détectable des membres, montrant ainsi que la perte d'activité de ZRS n'est pas responsable de la polydactylie préaxiale. Ces observations confirment que les mutations ponctuelles dans ZRS n'inactivent pas la voie Shh mais la redirigent vers un site ectopique.
D'autres anomalies génétiques suggèrent que des régulateurs additionnels sont impliqués dans la construction des membres. Actuellement la complexité de cette régulation n'est pas éclaircie puisque seul ZRS a été identifié. Puisque les transgènes induisent une expression ailleurs que dans le site normal, les mutations ponctuelles affectant ZRS apparaissent comme seules requises pour déterminer une expression ectopique.
Notre étude a montré que des mutations ponctuelles dans un cis-régulateur essentiel au développement peuvent se traduire par des éléments squelettiques additionnels, confirmant la notion que de petites modification de la régulation peuvent avoir de grandes incidences sur la morphologie de l'animal.
[1.] Le séquencage des génomes a montré que les séquences d'ADN étaient plus ou moins variables, donc susceptibles de mutations (du moins de leur conservation). Celles qui correspondent aux gènes contrôlant le développement la construction de l'organisme lors de la période embryonnaire, font partie de celles qui varient peu : On peut reconnaître leur similitude à travers toutes les espèces Invertébrées et Vertébrées. C'est le cas des gènes homéotiques ou gènes architectes, par exemple HoxA et HoxD intervenant dans la construction du membre et de sa partie distale, main ou pied. C'est aussi le cas des séquences régulatrices du fonctionnement des autres gènes.En fait, il apparaît logique que les séquences clés de processus complexes aient été conservées tout subissant et conservant certaines mutations : C'est ainsi que peuvent s'expliquer la diversité des formes de vie, et la variabilité affectant un plan de construction ancestral.
[2.] Les régulateurs cis sont de courtes séquences d'ADN positionnées à côté de certains gènes. Leur fonction est de contrôler l'expression d'autres gènes en jouant le jeu d'interrupteurs ou de potentiomètres sur ceux-ci.
[3.] Le gène Shh code pour une protéine Shh qui active une chaîne de réactions en cascade déclenchant l'expression de gènes architectes. Cette chaîne de réactions porte le nom (humoristique !) de voie sonic hedgehog. Elle intervient lors de la vie embryonnaire dans la construction des membres mais aussi dans celle du système nerveux, entre autres.
Quant au nom donné à ce gène ? Il faut croire en l'humour des biologistes !

[4.] Dans le génome humain, ZRS est positionné à proximité du gène Shh. Sa séquence longue d'environ 800 paires de bases est très conservée dans tous les génomes des Vertébrés, avec cependant des différences suivant les espèces. Elle est très ancienne, présente chez les premiers Vertébrés (au moins !) puisque son rôle dans le développement des appendices a été récemment identifié chez les Poissons Chondrichthyiens (Poissons «cartilagineux» type requin et raie).
[5.] A partir de la marge où le régulateur est activé, un gradient de concentration des acteurs moléculaires est crée suivant l'axe de polarité du membre. Ce sont des différences de concentration de certaines molécules qui expliquent la différence de forme des doigts qui sont construits donc l'asymétrie de la main et du pied : Pouce à 2 phalanges au pôle antérieur éloigné de la ZPA et petit doigt au plus proche de cette zone d'activation de ZRS.
[6.] Les plus célèbres des chats polydactyles sont les chats d'Hemingway, descendant tous d'un chat polydactyle, Snowball, qui fut donné à l'écrivain vers 1935.Cette colonie représentait une centaine de chats du vivant de l'auteur. Aujourd'hui, leurs descendants continuent à vivre dans la demeure de l'écrivain transformée en musée à Key West.
[7.] Tomoko Sagai, Masaki Hosoya, Youichi Mizushina, Masaru Tamura and Toshihiko Shiroishi.
Mammalian Genetics Laboratory, National Institute of Genetics, Yata-1111 Mishima, Shizuoka-ken 411-8540, Japan.
"Elimination of a long range cis-regulatory module cause complete loss of limb-specific Shh expression and truncation of the mouse limb". Etude en anglais téléchargeable ici (PDF).
Le mot d'Odile
«Comme ces mutations produisent chez l'Homme un phénotype spécifique aux membres, sans autre anomalie physiologique discernable, nous proposons le fait que ce type de polydactylie n'a pas d'incidence nuisible sur la santé du chat.»
Les générations successives de MC polydactyles ne montrent que cela : «Des éléments squelettiques supplémentaires», des doigts dans le cas de cette mutation, et rien que des doigts. Nulle autre «incidence nuisible»
… «il y a une relation entre une mutation spécifique et le phénotype observé». Il n'est pas relaté, par ceux qui ont élevé et élèvent toujours des MC poly, une variabilité du phénotype au delà du nombre de doigts supplémentaires et du fait que les quatre pattes peuvent être affectées comme seulement les deux antérieures.
La mutation portée et transmise par les MC n'est pas comparable à la mutation Cuban des humains ou Hs des souris, qui se traduisent par d'autres anomalies lourdes. Leur seul point commun est qu'elles sont aussi ponctuelles et situées sur la même séquence d'ADN.
La sélection d'élevage des polydactyles a veillé à ce qu'aucune autre mutation affectant la morphologie des membres ne soit introduite dans les lignées. Les mariages sont même maintenant autorisés entre deux polydactyles, leur descendance ne montrant toujours qu'un nombre surnuméraire de doigts. La colonie des chats d'Hemingway, construite à partir d'un seul ancêtre poly et qui a perpétué cette caractéristique dans le cadre d'une certaine consanguinité sans doute, illustre par l'absence d'autres anomalies l'aspect bénin de la mutation Hw que portent ausi les MC.
Si débat il y a, il ne doit plus se nourrir d'extrapolations sur les risques potentiels d'anomalies squelettiques et fonctionnelles graves. Les arguments pour interdire les polydactyles en exposition, en l'état actuel des connaissances, ne peuvent plus être d'ordre biologique. Sinon, bien d'autres races félines sont aussi à remettre en cause. Car où est la cohérence entre la reconnaissance totale d'une race comme le Munchkin (porteur d'une mutation qui raccourcit les pattes) et le refus des MC porteurs de doigts supplémentaires ? Où est la cohérence entre le fait d'admettre les MC poly en reproduction et celui de les refuser en exposition ?
Les arguments actuels ne peuvent plus être que du domaine de l'esthétique puisque les expos, ce n'est que cela… Qui oserait définir deux colonnes, celle du beau et celle du laid ?

Le Maine Coon polydactyle. Traduction de l'article de Susan Grindell. Chatterie Mainelymagic Maine Coons
La polydactylie. Traduction de l'article de Solveig Pflueger.
Observation sur la polydactylie. Résumé de l'article de Sheila Curtis.
Polycats Association Française du Chat Polydactyle - Loi 1901 -.